Väčšina mastných kyselín v organizme pochádza z potravy. Niektoré bunky ich však dokážu vytvárať – syntetizovať. Táto syntéza mastných kyselín prebieha hlavne v pečeni. Obmedzenú schopnosť syntézy majú adipocyty (bunky tukového tkaniva), mliečna žľaza tvoriaca mlieko, mozog a tenké črevo.
Hlavným substrátom je acetyl-CoA. Keď je ho v bunke dostatok a nie je potrebné ho využiť na tvorbu energie, začne syntéza mastných kyselín (po jedle, pri dostatku glukózy a aminokyselín – acetyl-CoA pochádza z glykolýzy, alebo metabolizmu aminokyselín).
Syntéza mastných kyselín
Na začiatok si rozdeľme celý proces do niekoľkých krokov.
1. Väčšina acetyl-CoA vzniká v matrix mitochondrie, zatiaľ čo enzýmy pre tvorbu mastných kyselín sa nachádzajú v cytoplazme. Substrát teda potrebujeme preniesť cez membránu do cytoplazmy bunky. Vo forme acetyl-CoA to nie je možné.
Vieme, že pri odbúravaní mastných kyselín sme na transport cez membránu využívali karnitín. V opačnom smere sa však používa iný spôsob. Acetyl-CoA vstúpi do citrátového cyklu.
Produktom prvej reakcie je citrát. Ak má bunka dosť energie, citrátový cyklus sa v tomto momente zastaví. Citrát sa hromadí a vstupuje do novej dráhy, syntézy mastných kyselín. Transportuje sa cez vnútornú mitochondriálnu membránu a dostáva sa do cytosolu.
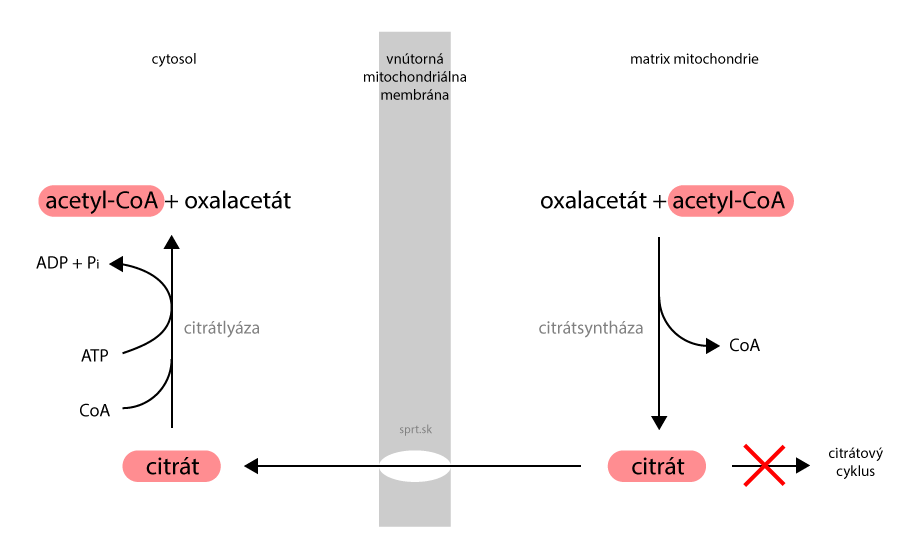
2. V cytoplazme máme v tomto momente citrát, no syntéza mastných kyselín vyžaduje inú molekulu. Citrát sa rozloží na pôvodné produkty, oxalacetát a acetyl-CoA. Spotrebuje sa pri tom energia vo forme 1 ATP.
Čo sa deje s oxalacetátom?
Oxalacetát je v cytoplazme redukovaný na malát (posledná reakcia citrátového cyklu v opačnom smere). Ten z časti prechádza naspäť do matrix mitochondrie. Časť malátu je pomocou tzv. jablčného enzýmu premenená na pyruvát. Táto cesta je výhodnejšia, pretože vzniká NADPH + H+ (v reakcii oxalacetát → malát vzniká NAD+). Syntéza mastných kyselín vyžaduje dostatok NADPH + H+.
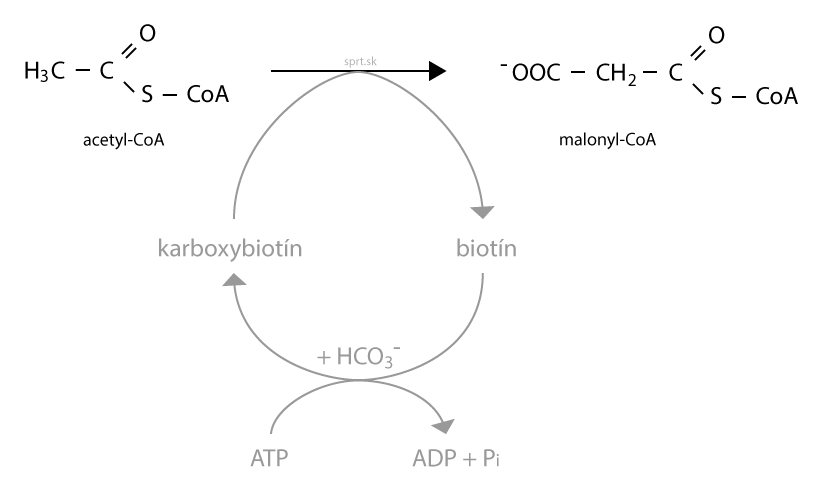
Ešte však nie je koniec. Acetyl-CoA nemá dostatok energie na to, aby priamo vstúpil do ďalšej reakcie. Musí sa aktivovať, čím označujeme jeho karboxyláciu. Výsledným produktom je malonyl-CoA.
Nasledujúci obrázok ukazuje reakcie, ktoré vedú k tvorbe malonyl-CoA. Acetyl-CoA nedokáže na seba naviazať CO2, potrebný je kofaktor. Najčastejším kofaktorom pri karboxylačných reakciách je biotín. Ten sa v prvom kroku karboxyluje, pri čom spotrebuje ATP. Vzniká karboxybiotín, z ktorého sa karboxylová skupina prenáša na acetyl-CoA. Táto reakcia je regulačná, určuje rýchlosť syntézy mastných kyselín.
3. Malonyl-CoA sa pripája k enzýmom, ktoré katalyzujú tvorbu mastných kyselín. Výsledným produktom je mastná kyselina.
Syntéza mastných kyselín na enzýmovom komplexe
Syntéza mastných kyselín prebieha na komplexe tvorenom až 7 enzýmami. Preto sa nazýva multienzýmový komplex – syntháza mastných kyselín. Okrem katalytických častí tento komplex obsahuje aj tzv. ACP – acyl carrier protein – časť, ktorá viaže 4-fosfopanteteín.
Fosfopanteteín je derivát kyseliny pantoténovej (vitamín B5). Jeho úlohou je prostredníctvom –SH skupiny viazať medziprodukty vznikajúce počas syntézy mastných kyselín. Dá sa povedať, že nahrádza koenzým A a plní jeho úlohu.
Acyl, ktorý vzniká pri syntéze mastných kyselín, sa teda viaže na –SH skupinu na konci reťazca fosfopanteteínu.
Syntéza mastných kyselín, reakcia po reakcii
Ako sme uviedli vyššie, reakcie prebiehajú na enzýmovom komplexe nazývanom syntháza mastných kyselín v prípade, že bunka má dostatok energie, glukózy a aminokyselín. Substrátom pre tvorbu je acetyl-CoA, ktorý sa transportuje z matrix mitochondrie do cytosolu a následne dôjde k jeho aktivácii. Vzniká malonyl-CoA.
Naviazanie substrátov
Máme dva potrebné substráty: malonyl-CoA a acetyl-CoA. Oba sa viažu na –SH skupiny proteínov v multienzýmovom komplexe.
Acetyl-CoA sa prenáša na –SH koniec enzýmu s názvom acetyl-CoA-transacyláza.
Malonyl-CoA sa prenáša na –SH koniec fosfopanteteínu.
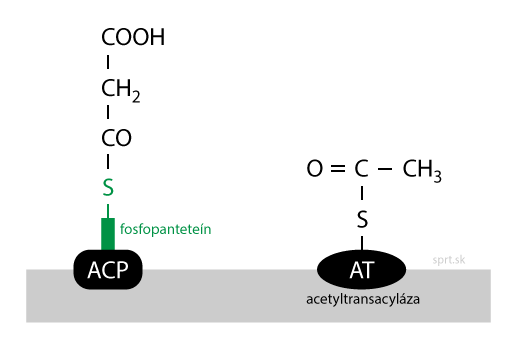
Kondenzácia
V tomto kroku dochádza k spojeniu dvoch substrátov. Acetylová skupina sa prenáša na malonylovú, vzniká oxoacyl. Pri reakcii dochádza k dekarboxylácii, odštepuje sa CO2 z –COOH (karboxylovej) skupiny na konci malonylového reťazca.
Oxoacyl, ktorý je produktom, je viazaný na fosfopanteteín prostredníctvom thioesterovej väzby (cez –SH skupinu). Reakciu katalyzuje enzým 3-oxoacylsyntháza (OAS).
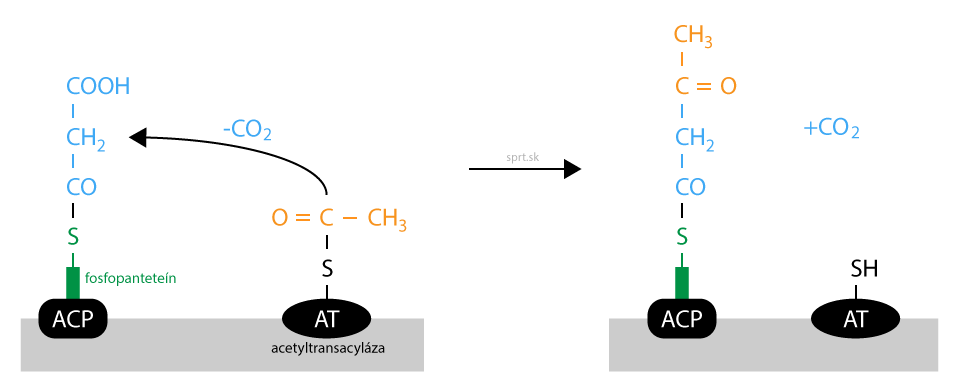
Redukcia
Oxoacyl viazaný na fosfopanteteín podlieha v ďalšom kroku redukcii. Po nej nasledujú ďalšie deje, ktorých spoločným cieľom je odstrániť oxoskupinu na treťom uhlíku.
Potrebný je kofaktor NADPH + H+, ktorý sa oxiduje na NADP+. Vzniká hydroxyacyl, stále viazaný na fosfopanteteín. Enzýmom katalyzujúcim reakciu je 3-oxoacylreduktáza.
Všimnite si, že reakcia je hydrogenáciou. Pri odbúravaní, β-oxidácii mastných kyselín, je prvou reakciou dehydrogenácia. Aj nasledujúce deje sú pri syntéze opačné, ako pri odbúravaní.
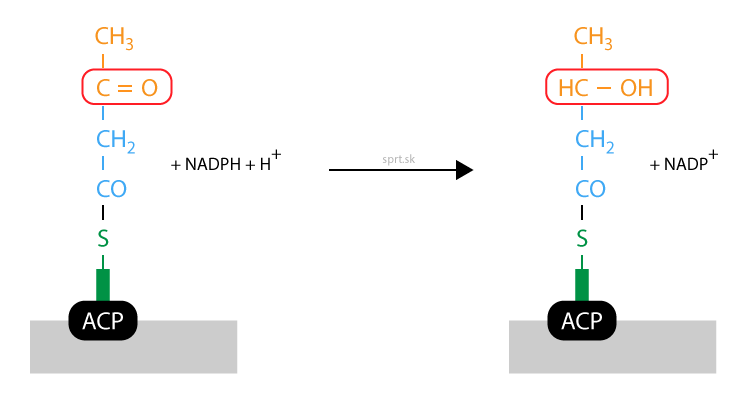
Dehydratácia
Enzým 3-hydroxyacyldehydratáza katalyzuje ďalšiu z reakcií. Z hydroxyacylu sa odštepuje voda a medzi druhým a tretím uhlíkom vzniká dvojitá väzba. Produkt sa nazýva trans-2-enoyl, stále je viazaný na fosfopanteteín.
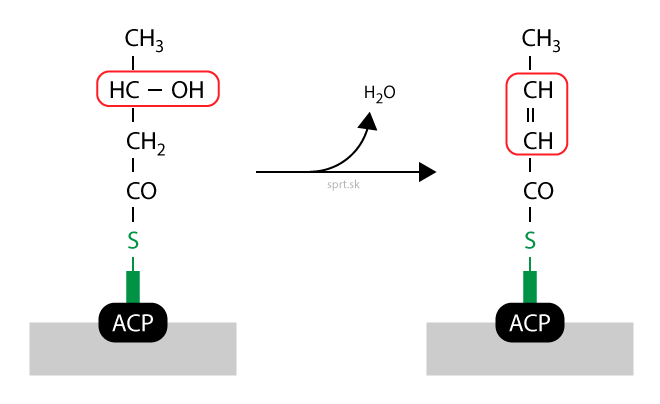
Redukcia
Dvojitá väzba zaniká vďaka redukcii – hydrogenácii – a vzniká nasýtený acyl. Ten je stále viazaný na rovnakom mieste, –SH skupine fosfopanteteínu. Reakciu katalyzuje enoylreduktáza a potrebný je kofaktor NADPH + H+.
Ako vidíte na obrázku, acyl má v tomto momente štyri atómy uhlíka. Acetyl-CoA sme teda predĺžili o dva uhlíky.
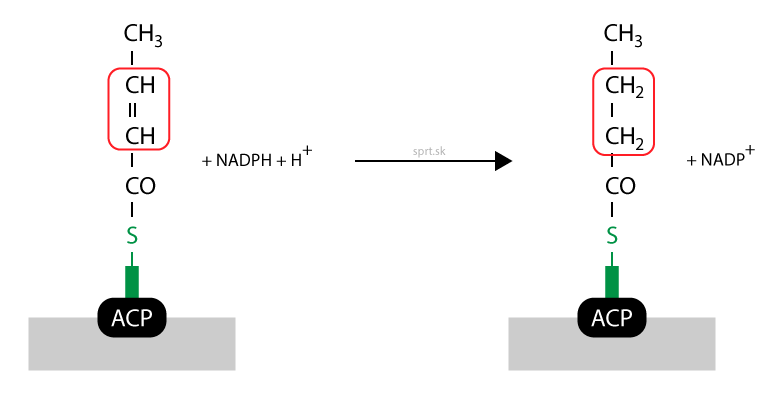
Syntéza mastných kyselín
Novovytvorený acyl sa prenesie z fosfopanteteínu na oxoacylsyntházu, aby mohla syntéza ďalej pokračovať. Uvoľní sa tak –SH skupina fosfopanteteínu, na ktorú sa naviaže ďalší malonyl-CoA. Ten kondenzuje s „naším“ acylom a vyššie uvedené reakcie prebiehajú opäť. Výsledkom je nasýtený acyl predĺžený o ďalšie dva uhlíky.
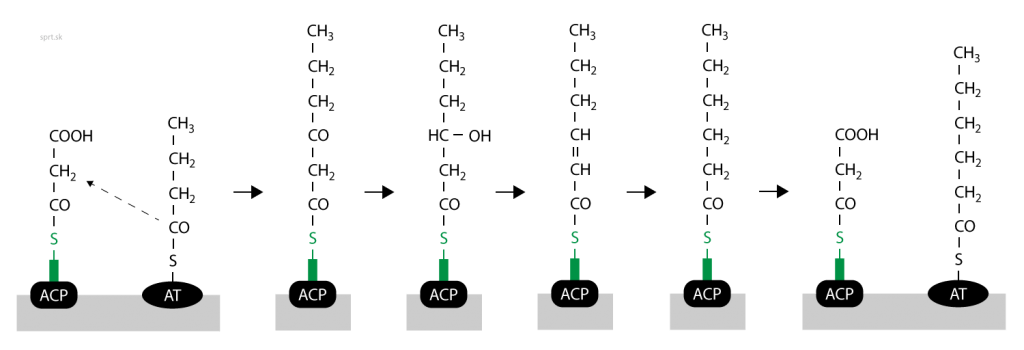
Keď všetky tieto kroky opakujeme niekoľkokrát, výsledkom sú mastné kyseliny. Najčastejším produktom je kyselina palmitová, ktorá má 16 uhlíkov. V menšom množstve sa produkuje aj kyselina steárová, 18-uhlíkatý produkt.
Výsledkom reakcií je nasýtený acyl s daným počtom uhlíkov naviazaný na fosfopanteteín. Pomocou thioesterázy sa hydrolyticky odštepuje palmitát, respektíve stearát. Na konci fosfopanteteínu sa obnovuje –SH skupina.
Energetická bilancia
Určime si energetickú bilanciu syntézy kyseliny palmitovej. Má 16 uhlíkov, takže na jej tvorbu sa použije 8 acetyl-CoA a sedemkrát prebehne vyššie popísaný cyklus reakcií.
Z ôsmich acetyl-CoA sa sedem premieňa na malonyl-CoA. Na každú z týchto reakcií spotrebujeme 1 ATP, takže spolu 7 ATP.
V každom cykle reakcií sú následne dve redukcie. Pri nich sa spotrebuje NADPH + H+. Povedali sme, že pri 16-uhlíkatej kyseline prebehne tento cyklus sedemkrát. To znamená, že použijeme 7×2=14 NADPH + H+.
Zhrnutie
Acylový zvyšok dosiahne dĺžku reťazca 16 uhlíkov po siedmich cykloch. Následne sa hydrolyticky odštiepi palmitát, hlavný produkt syntézy mastných kyselín.
Actetyl-CoA má dva uhlíky, takže na tvorbu 16-uhlíkatého reťazca spotrebujeme 16:2=8 molekúl acetyl-CoA. Na začiatku syntézy sa acetyl-CoA karboxyluje na malonyl-CoA za spotreby 1 ATP a 1 CO2, ktorý vstupuje do ďalších reakcií. Viaže sa na fosfopanteteín. Druhým substrátom je acetyl-CoA, pripevnený na enzým acetyl-CoA-transacyláza.
Acetyl-CoA a malonyl-CoA kondenzujú a vo väzbe na fosfopanteteín prebieha sled reakcií, pri ktorých spotrebujeme 2 NADPH + H+. Výsledkom je štvoruhlíkatý acyl pripevnený na fosfopanteteín. V ďalšom kroku sa acyl prenesie na enzým oxoacylsyntháza.
Ďalšia molekula acetyl-CoA podlieha karboxylácii a malonyl-CoA sa viaže na fosfopanteteín. Opäť dochádza ku kondenzácii a nasledujúcim reakciám – spotrebovalo sa teda 1 ATP na karboxyláciu, 1 CO2 a 2 NADPH + H+.
Toto sa opakuje až do ukončenia syntézy. Na každé predĺženie reťazca o dva uhlíky teda potrebujeme CO2, ATP a 2 NADPH + H+. V konečnom dôsledku sa tak spotrebuje 1 acetyl-CoA, 7 malonyl-CoA (z acetyl-CoA), 7 ATP, 7 CO2, 14 NADPH + H+ a 6 H2O na odštiepenie acylu z fosfopanteteínu.
Ako vidíte, syntéza mastných kyselín je energeticky veľmi náročný proces, ktorý zároveň vyžaduje množstvo dostupného acetyl-CoA.
Odkiaľ bunka získava NADPH + H+?
Najväčším producentom je pentózafosfátová dráha. Glukóza sa v nej premieňa na päťuhlíkaté cukry a produkuje sa NADPH + H+. To bunka využíva pri redukčných syntézach, príkladom je práve syntéza mastných kyselín.
Druhou možnosťou je dekarboxylácia malátu pomocou „jablčného“ enzýmu. Spomeňte si, že sme o tejto dráhe hovorili už vyššie v texte. Pri transporte acetyl-CoA z mitochondrie do cytosolu sa do cytosolu dostane oxalacetát. Ten sa redukuje na malát a ten má dve možnosti – transport naspäť do matrix, alebo dekarboxylácia sprostredkovaná jablčným enzýmom. Pri tom vzniká pyruvát, ktorý sa transportuje do matrix mitochondrie.
Regulácia syntézy
Syntéza mastných kyselín je regulovaná na troch úrovniach.
1.) Bude prebiehať len v prípade, že v cytosole je dostatočná dostupnosť substrátov – acetyl-CoA a NADPH + H+.
2.) Dôležitá je aj allosterická regulácia, t.j. na úrovni celej bunky. Tu majú hlavné slovo dva enzýmy: izocitrátdehydrogenáza a acetyl-CoA-karboxyláza.
Izocitrátdehydrogenáza je enzým citrátového cyklu, ktorý katalyzuje nevratnú reakciu izocitrát → 2-oxoglutarát. To znamená, že aby prebiehala syntéza mastných kyselín, tento enzým musí byť inhibovaný. Inhibíciu spôsobujú molekuly ATP a NADH + H+, ktoré informujú bunku o dostatku energie. Citrátový cyklus sa zablokuje a hromadiaci sa citrát prestupuje do cytoplazmy, kde sa následne rozloží na acetyl-CoA potrebný na syntézu MK a oxalacetát.
Acetyl-CoA-karboxyláza katalyzuje premenu acetyl-CoA na malonyl-CoA, takže ide o nevyhnutnú reakciu pre syntézu mastných kyselín. Enzým musí byť preto aktivovaný. Aktivuje sa v dôsledku zvýšeného množstva citrátu v cytosole. Naopak, inhibuje ho palmitoyl-CoA, ktorý signalizuje odbúravanie mastných kyselín – a syntéza s odbúravaním nemôžu prebiehať súčasne.
3.) Komunikácia medzi bunkami je ďalšia úroveň regulácie, zabezpečujú ju hormóny. Enzým acetyl-CoA-karboxyláza je aktivovaný inzulínom, zatiaľ čo glukagón a adrenalín ho inhibujú.
Toto si viete jednoducho odvodiť. Inzulín sa produkuje po jedle a podporuje vychytávanie glukózy z krvi. Tá preniká do buniek a v zvýšenej miere dochádza ku glykolýze. Produktom je pyruvát, ktorý sa premieňa na acetyl-CoA. Keďže ho je nadbytok a vďaka glykolýze máme dostatok energie, presúva sa acetyl-CoA do cytosolu. Tam sa využije na tvorbu mastných kyselín. Zjednodušene môžeme povedať, že energiu a živiny z potravy telo ukladá do tukových zásob (mastné kyseliny sa neskôr zabudujú do triacylglycerolu).
Glukagón je hormón vylučovaný pri hladovaní, takže má opačné účinky. Podporuje odbúravanie tukov, aby sme získali energiu a produkty využiteľné na syntézu glukózy.
Adrenalín sa vylučuje v strese – fyzickom, aj psychickom. Úlohou je pripraviť telo na boj alebo útok. Na to potrebujeme akumulovať množstvo energie pre svaly, takže dôjde k odbúravaniu mastných kyselín. Syntéza je energeticky náročná, preto prebiehať nemôže.
Predĺženie reťazca a dvojité väzby
Produktom syntézy mastných kyselín je predovšetkým kyselina palmitová, v menšej miere steárová. Iba tieto dve však organizmu nestačia. Preto máme systémy na predĺženie ich reťazca a tvorbu nenasýtených mastných kyselín. Tieto reakcie prebiehajú v endoplazmatickom retikule.
Mastná kyselina sa transportuje do endoplazmatického retikula a dôjde k naviazaniu koenzýmu A. Predlžovanie, elongácia, následne prebieha podobne ako v cytosole – pomocou malonyl-CoA. Enzýmy však nie sú spojené v komplexe.
Desaturácia - tvorba dvojitých väzieb
Tento proces prebieha takisto v endoplazmatickom retikule a výsledným produktom sú nenasýtené mastné kyseliny.
Na endoplazmatickom retikule buniek pečene sú viazané proteíny – enzýmy desaturázy. U človeka rozoznávame Δ9 (delta 9), Δ6, Δ5 a Δ4 desaturázy, rastliny majú navyše Δ12 a Δ15 desaturázy.
Podľa pomenovania vieme, kde vzniká dvojitá väzba. Napríklad Δ9 desaturáza vytvára dvojitú väzbu medzi 9. a 10. uhlíkom (počítame od –COOH konca).
Pri desaturácii mastných kyselín s dlhým reťazcom je prvým krokom práve Δ9 desaturácia. U živočíchov môže nasledovať tvorba dvojitej väzby už len medzi prvým a deviatym uhlíkom. Rastliny majú však vďaka ďalším desaturázam možnosť tvorby dvojitej väzby aj medzi deviatym a posledným uhlíkom reťazca. To je dôvod, prečo sú niektoré mastné kyseliny pre nás esenciálne – nemáme enzýmovú výbavu na to, aby sme ich vytvorili, jedinou možnosťou je prijať ich v potrave. Konkrétne ide o kyselinu linolovú a α-linolénovú.
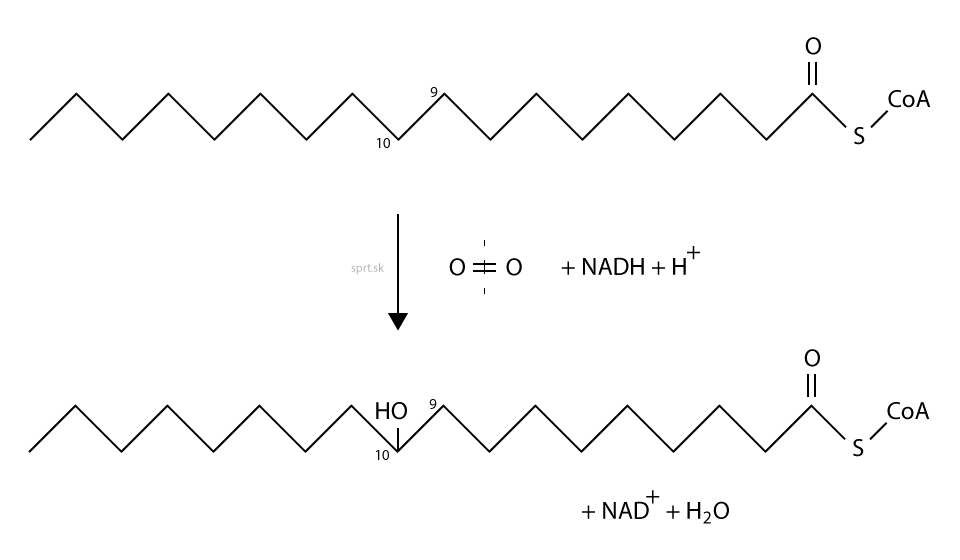
Priebeh desaturácie
Reakcia prebieha v dvoch krokoch v endoplazmatickom retikule.
Najprv sa mastná kyselina hydroxyluje. Ide o mechanizmus monooxygenázovej reakcie, potrebujeme molekulárny kyslík O2. Jeden atóm kyslíku sa zabuduje do –OH skupiny, druhý sa stane súčasťou vody. Okrem kyslíka teda potrebujeme aj NADH + H+.
Hydroxyacyl sa následne dehydratuje, t.j. dochádza k eliminácii vody a tvorbe dvojitej väzby. Výsledná konfigurácia je vždy vo forme cis.
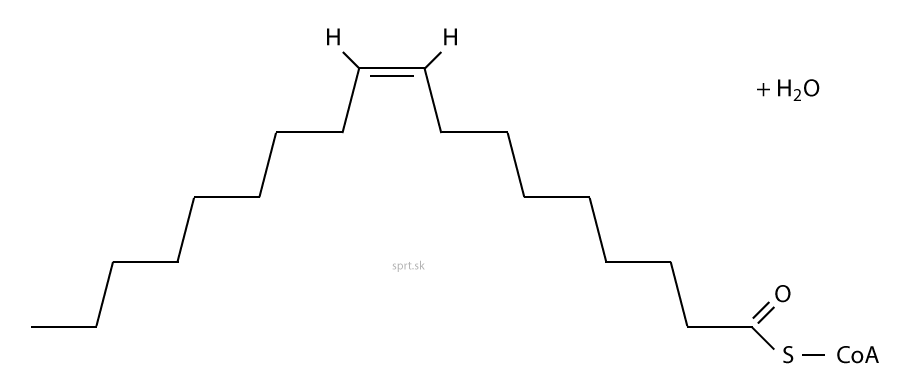
Esenciálne mastné kyseliny
Už sme si povedali, že esenciálne mastné kyseliny sú dve – linolová a α-linolénová. Ich tvorba vyžaduje prítomnosť desaturáz, ktoré v našom organizme nenájdeme. Musíme ich preto prijímať potravou.
Ak ich však prijmeme, dokážeme ich ďalej modifikovať prostredníctvom elongácie a desaturácie. Organizmus tak môže vyrobiť napríklad kyselinu arachidonovú, eikosapentaenovú a ďalšie. Ich predlžovaním a tvorbou dvojitých väzieb vznikajú polynenasýtené mastné kyseliny.
Zdroje:
Zdroje: prezentácia Lipidy II (tenzidy, trávení lipidů, mastné kyseliny – syntéza a odbourání) Biochemického ústavu Lekárskej fakulty Masarykovej univerzity (2017)
https://en.wikipedia.org/wiki/Fatty_acid_desaturase